

Tran Van Tien1, Nguyen Hoang Nghia2 and Nianhe Xia3
1Da Lat University
2Vietnamese Academy of Forest Sciences
3South China Botanical Garden
. INTRODUCTION
The usefulness of anatomical characters of the epidermis of bamboos in classification has been recognized for a long time. Brandis (1907) studied the structure of bamboo leaves with special reference to their upper (adaxial) and lower (abaxial) epidermis. Takenouchi (1941) published detailed account of Japanese bamboos with special reference to their morphology, anatomy and development.
The shape of epidermal cells and stomata are also characteristic and can often be of value for diagnostic purposes. Pattanath and Rao (1969) evaluated the importance of anatomical features in the identification of bamboos. They clearly show that the epidermal features arrange along with internodal structure and is very useful in differentiating them. Furthermore, several authors (Wu, 1962; Yang et al., 2008) have also studied the shape of stomata of Schizostachyum species from China. The detailed studies show that the shape of stomata on the lower epidermis covers finger – like protuberances (papillae). They come forth from the epidermal cells next to the epidermal cells and next to the guard cells. They are obvious variations in papillae forms and distributional patterns around the stomatal apparatus amongst the examined genera and species. Their significance is indicating taxonomic value.
The aims of this investigation are to provide the basis for an authoritative description of the anatomical structure of certain, to determine the variability an possible trends the stomata on the lower epidermis covers finger – like protuberances and hairs – microhairs between leaf blades of different species, which could be used in an identification for the Schizostachyum species from Vietnam.
II. MATERIALS AND METHODS
The samples of this study were collected from wild populations. The origins of the material used in the present study are given in table 1. The materials for scanning electron microscopy (SEM) were submerged in xylene for at least 4 hous in order to remove the waxy covering from the leaf epidermis, and then were attached directly to Petri dish without any treatment. After gold sputtering, the specimens were examined and photographed. The terminology of epidermis appendages follows Metcalfe (1960), Wu (1962) and Yang et al., (2008).
Table 1. Taxa and voucher for species sequenced in this study
| Taxa | Woucher No. | Source |
|
Schizostachyum hainanense |
1220080467 (FSIV) |
Hue, Vietnam |
|
Schizostachyum sp1 |
1020100471 (FSIV) |
Baria, Vietnam |
|
Schizostachyum pseudolima |
1220080465(FSIV) |
Phu Tho, Vietnam |
|
Schizostachyum funghomii |
1220080464(FSIV) |
Phu Tho Vietnam |
|
Schizostachyum nghianum |
1200904712(FSIV) |
Tuyen Quang, Vietnam |
|
Schizostachyum aciculare |
0820090466(FSIV) |
Phu Quoc Island, Vietnam |
|
Schizostachyum mekongense |
1200904714(FSIV) |
Lam Dong, Vienam |
|
Schizostachyum sp2 |
20090470 (FSIV) |
Lam Dong, Vietnam |
|
Schizostachyum sp3 |
1520090466 (FSIV) |
Hai Duong, Vietnam |
|
Schizostachyum yalyense |
06200504662 (FSIV) |
Gia Lai, Vietnam |
|
Schizostachyum langbianense |
1020060460 (FSIV) |
Lam Dong, Vietnam |
|
Schizostachyum ninhthuanense |
1512200804661 (FSIV) |
Ninh Thuan, Vietnam |
|
Schizostachyum dullooa |
0710600 (FSIV) |
Tuyen Quang, Vietnam |
III. RESULTS
The foliage leaf epidermis of 13 species of Schizostachyum investigated (fig. 1 – 5). It seems to be made up of the forms of short cells, long cells, midrib of the leaf blade and silica bodies are more conspicuous when examined with SEM similar to the results of previous studies (Metcalfe, 1956, 1960; Tateoka, 1959; Wu, 1962; Clark, Londono, 1991, Chen et al., 1992). In case of the genus Schizostachyum, epidermal cells are highly modified and become quite different in appearance from the original cell. These characters are important in classification such as: hairs – microhairs and palliate forms; and distributional patterns around the stomata apparagus of the abaxial and adaxial foliage leaf epidermis (Wu, 1962; Yang et al., 2008). They are presented in table 2 and figure: 1 – 5.
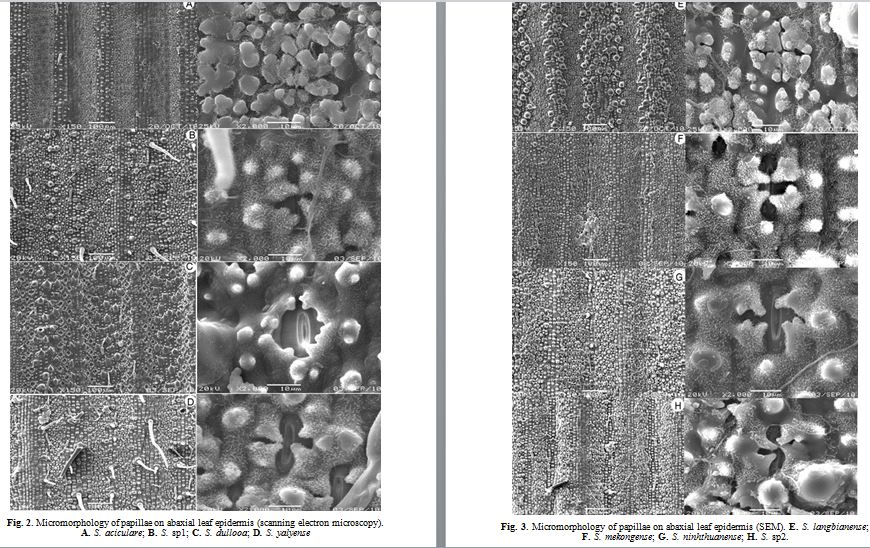
Table 2. Characters of foliage leaf epidermis (SEM)
| Name of species | Hairs | Stomata | Figure | ||||
| B | S | G | L | Rows | Papillae forms and distributional patterns around the stomatal apparatus |
||
|
S. aciculare |
+ |
4, triangular, overarching and surrounded |
A | ||||
|
S. sp2 |
+ | 4 – 5 |
4, triangular, overarching and long |
H | |||
|
S. sp3 |
+ | 4 – 5 |
4, triangular, overarching and long |
I | |||
|
S. sp1 |
+ | + | 2 – 4 |
4, triangular, overarching and long |
B | ||
|
S. dullooa |
+ | + | 4 – 4 |
4, triangular, overarching and short |
C | ||
|
S. funghomii |
++ | + | 4 – 4 |
4, triangular, overarching and surrounded |
M | ||
|
S. nghianum |
+ | 4 – 4 |
4, triangular, overarching and surrounded |
L | |||
|
S. hainanense |
++ | + | + | 2 – 4 |
4, triangular, overarching and long |
K | |
|
S. langbianense |
++ | + | 4 – 4 |
4, triangular, overarching and surrounded |
E | ||
|
S. ninhthuanense |
++ | + | + | 4 – 5 |
4, triangular, overarching and surrounded |
G | |
|
S. pseudolima |
++ | + | 2 – 4 |
4, triangular, overarching and long |
J | ||
|
S. mekongensis |
++ | + | 4 – 5 |
4, triangular, overarching and long |
F | ||
|
S. yalyense |
+ | ++ | ++ | 4 – 5 |
4, triangular, overarching and surrounded |
D | |
3.1. Hairs – Microhairs
Hairs – Microhairs commonly seen and consist of three types such as long hairs, geniculate hairs, and spines:
3.1.1. Long hairs (L)
Long hairs are narrow, long and one celled hairs. They are usually found on the epidermis covers but widely distributed over the completely lower epidermis of some species (S. funghomii, S. nghianum, S. hainanense, C. langbianense, S. ninhthuanense and S. yalyense). However, long hairs are abundant in S. yalyense.
3.1.2. Geniculate hairs (G) (bicellular hairs)
Geniculate hairs arise from the derivatives of the short epidermal cells (Wu, 1962), which are usually broken off by external forces and distributed on the lower epidermis covering the sclerenchyma region. They situated on the lower epidermis of some species (S. sp2, S. sp3, S. hainanense, S. langbianense, S. ninhthuanense, S. pseudolima, s. mekongense and S. yalyense). However, long hairs are abundant in S. yalyense.
3.1.3. Spines hairs (S)
Spines hairs are short, spine – like cells, which are rounded at the base and acute at the apex (Wu, 1962). They are usually dense present on both surfaces over sclerenchyma region in some species (S. hainanense, S. langbianense, S. ninhthuanense, S. mekongense), but sparse in S. yalyense, S. funghomii and S. pseudolima.
3.2. The stomata and papillae
The stomata of all species on the lower epidermis are larger, elliptical in shape and arranged in 2 – 5 rows on each side of a vein. Each stoma covers finger – like protuberances (papillae) which come forth from the epidermal cells next to the epidermal cells, incline towards the center of stoma (Wu, 1962). There are obvious variations in papillae forms and distributional patterns around the stomatal apparatus amongst the examined genera, and their significance is to indicate taxonomic value (Wu, 1962; Yang et al., 2008). There are usually four triangular papillae overarching each individual stomata apparagus in all species, but otherwise, S. langbianense, S. aciculare, characterized by having four triangular papillae overarching each individual stomata apparatus and surrounded. However, four triangular papillae overarching each individual stomata apparatus are unequal in length amongst species. There are two types:
– The triangular papillae overarching are surrounded, which found in S. dullooa, S. nghianum, S. ninhthuanense and S. yalyense;
– The triangular papillae overarching are long, which found in some other species.
In addition to the papillae, the number of rows of stomata is also worth noting (Wu, 1962). On the lower epidermis in Schizostachyum, the number of rows of stoma varies from 2 to 5 rows on each side of a vein and depends on the species. The number rows of stoma can be divided into 4 patterns:
In some case, there are 4 – 5 rows found in S. yalyense and S. ninhthuanense.
In some case, there are 4 – 5 rows found in S. sp2, S. sp3 and S. mekongense.
In some case, there are 3 – 4 rows found in S. funghomii, S. sp2 and S. langbianense.
In some case there are 2 – 3 rows found in S. hainanense, S. pseudolima.
3.3. Discussion and conclusion
Wu (1962) shown about the variation in occurrence and distribution of microhairs. It is important to know whether the walls of the basal cells of microhairs are smooth. The leaf epidermal characters of Schizostachyum species in general, and the papillae form and distributional pattern on the adaxial foliage leaf epidermis have been proven to be of
taxonomic value at the specific, generic (Metcalfe, 1960, Yang et al., 2008). In the current study, epidermal features were again constant within species. Schizostachyum species were confirmed that each individual stomatal apparatus is overarched by four branched of unbranched triangular papillae (Yang et al., 2008). However, four triangular papillae overarching each individual stomata apparatus are unequal in length amongst species, as discussed above.
These are important in classification such as hairs – microhairs and palliate forms and distributional patterns around the stomata of the abaxial foliage leaf epidermis. Thus, with the possible combination of several vegetative and anatomical characteristics, an experienced anatomist could succeed in identifying the species of Schizostachyum.
Acknowledgements
The authors would like to express their sincere thanks to Vietnamese Academy of Forest Science and Foundation of China (grant no. 30770155), South China Botanical Garden for supporting collected specimens and the investigating scanning electron microscopy.
REFERENCES
- Brandis, D., 1907. Remarks on the structure of bamboo leaves, Transactions of the Linnean Society of London, Botany. 7(5): 69 – 92.
- Chen, S.L., Jin, Y.X. and Wu, Z.J., 1993. Micromorphological atlas of leaf epidermis in Gramineae. Nanjing: Jiangsu Science and Technology Publishing House, 1 – 56.
- Clark, L.G. and Londono, X., 1991. A new species and new sections of Rhipidocladum (Poaceae: Bambusoideae). American Journal of Botany, 78: 1260 – 1279.
- Metcalfe, C.R., 1956. Some thoughts on the structure of bamboo leaves. Botanical Magazine, Tokyo. 69: 391 – 400.
- Metcalfe, C.R., 1960. Anatomy of the Monocotyledons I: Gramineae. London. Oxford University Press. ixi: 731 pp.
- Pattanath, P.G and Rao K.R., 1969. Epidermal and Internodel structure of the culm as an aid to identification and classification of bamboo. In: Recent advances in the Anatomy of Tropical seed plants, Hindustan Publishing Corporation, Delhi. 7: 179 – 196.
- Tateoka, T., Inowe, S. & Kawano, K., 1959, Notes on some grasses IX: Systematic significance of bicellular microhairs of leaf epidermis. Bot. Gaz. 121(2): 80 – 91.
- Takenouchi, Y., 1931. Systematisch – vergleichende Morphologie und Anatomie der Vegetationsorgane der japanischen Bambus – Arten. Mem. Faculty Science Agriculture, Taihoku Imp. Univ. III (1): 1 – 60.
- Wu, M.C.Y., 1962. Classification of Bambuseae based on leaf anatomy. Botanical Bulletin of Academia Sinica. 3: 83 – 107.
- Yang, H.Q., Wang, H. and Li, D.Z., 2008. Comparative morphology of the foliage leaf epidermis with emphasis on papillae characters, in key taxa of woody bamboos of the Asian tropics (Poaceae: Bambusoideae). Botanical Jounal Linnean Society. 156(3): 411 – 423.
Latest news
- Project: Study on integrated management of insect pests and diseases on Docynia indica in northern Vietnam
- Project: Study on integrated pest management of major leaf insect pests and stem borer beetles in Acacia hybrid, A. mangium and A. auriculiformis in Vietnam
- Project: Research and develop an integrated management program for control of cinnamon pests in some main cinnamon growing areas (Northern mountainous region and Quang Nam province)
- Project: Pilot plantation and completion of planting techniques of technologically advanced germplasm of Acacia auriculiformis (AA42, AA53, AA56, AA92, AA95) for sawlog in the southeast and south central.
- Project "Research on selection of varieties and intensive afforestation techniques of Prunus arborea (Blume) Kalkman for providing lager timber in the Northeast and Northwest regions, Vietnam"
Oldest news
- A new species od Adinandra jack. (Theaceae) from Vietnam
- Mapping the potential for REDD+ to deliver biodiversity conservation in Viet Nam
- Trajectories of deforestation, coffee expansion and displacement of shifting cultivation in the Central Highlands of Vietnam
- Progress in the genetic improvement for some main commercial planting tree species in Vietnam
- Research on the scientific basis for grouping wood species in Vietnam



